全基因组关联研究揭示了脑淋巴活动的机制
目录
1.引言
2.结果
2.1 ALPS指数的提取与评估
2.2 ALPS指数的遗传发现
2.3 全基因组关联分析结果
2.4 分层连锁不平衡评分回归(LDSC)分析
2.5 ALPS指数在全生命周期的遗传结构
2.6 ALPS指数在全生命周期的遗传结构
2.7 独立显著变异的遗传效应
2.8 独立显著变异的纵向验证
2.9 与神经精神表型和血管危险因素的遗传关联
2.10 ALPS指数与脑结构、白质完整性和脑功能的遗传相关性
2.11 ALPS指数的多基因结构及与神经精神表型和血管危险因素的遗传重叠
2.12 双向Mendelian randomization分析
2.13 基因水平分析与功能富集
2.14 细胞类型富集分析与功能分析
2.15 基因 - 药物相互作用分析
2.16 基因表达模式分析
3.讨论
3.1 12q23.3位点的强证据
3.2 ALPS指数的纵向变化与年龄依赖性遗传效应
3.3年龄依赖性关联与基因功能
3.4 ALPS指数的遗传结构与左右半球差异
3.5 胶质淋巴清除活动的遗传决定因素
3.6 ALPS指数、脑脊液tau病理和脑室体积的共同遗传机制
3.7 ALPS指数与脑结构及功能特征的遗传相关性
3.8 ALPS指数与ADHD风险的因果关系
3.9 研究局限性
4.总结
5.方法
5.1 研究被试
5.2 基因分型与质量控制
5.3 弥散张量成像(DTI)和ALPS指数计算
5.4 ALPS指数的可重复性
5.5 遗传力估计和全基因组关联分析
5.6 验证和泛化
5.7 元回归分析
5.8 纵向变化的GWAS
5.9 与其他表型的共享遗传变异
简要介绍:本研究通过全基因组关联分析(GWAS)揭示了脑类淋巴系统活动的遗传基础。研究发现17个全基因组显著位点和161个候选基因(如NUAK1、SULF2),其中部分位点(如2p23.3)在年轻与老年群体中呈现一致效应,提示其跨生命周期的调控作用。遗传相关性分析表明,ALPS指数与脑室体积、脑脊液tau蛋白水平及神经退行性疾病标志物存在关联,并发现与精神疾病(如ADHD)的多基因重叠。孟德尔随机化分析显示ALPS指数升高可降低ADHD风险(OR=0.647)。功能注释显示候选基因富集于神经发生、胶质细胞发育及氧化还原酶活性等通路。研究揭示了类淋巴系统遗传结构的生命周期特征及其与神经精神表型的关联,为探索类淋巴清除机制的生物学基础及疾病干预提供了新线索。
摘要:脑胶质淋巴活动,通过沿血管周围空间(ALPS)指数的弥散张量成像分析来指示,与发育性神经精神疾病和神经退行性疾病有关,但其遗传结构尚不清楚。在此,研究人员在英国生物样本库的31,021名个体的发现样本中,确定了17个独特的全基因组显著位点和161个与ALPS指数相关的候选基因。其中7个位点在两个独立的数据集中得到了验证。位于2p23.3位点的遗传信号在年轻和老年队列中均显示出显著的一致效应。遗传相关性和多基因重叠分析表明,ALPS指数、脑室体积和脑脊液tau水平之间存在共同的潜在遗传机制,GMNC(3q28)和C16orf95(16q24.2)是共同的遗传基础。研究的发现加深了对ALPS指数遗传学的理解,并为进一步研究胶质淋巴清除活动在生命周期内的神经生物学机制及其与神经精神表型的关系提供了见解。
1.引言
脑胶质淋巴系统是脑脊液(CSF)和脑组织间液交换的关键液体引流通道。它通过调节脑内液体和清除代谢废物(如β-淀粉样蛋白[Aβ]和tau)在维持脑稳态中发挥关键作用,并可能参与脑免疫功能。上述过程被认为受睡眠和昼夜节律的调节,这对于儿童和青少年早期大脑发育和神经认知功能至关重要。脑胶质淋巴功能障碍与发育性神经精神障碍有关,如注意力缺陷多动障碍(ADHD)和自闭症谱系障碍(ASD)。此外,越来越多的证据表明,脑胶质淋巴功能衰竭是神经退行性疾病(如阿尔茨海默病)的重要病理机制。
鉴于脑胶质淋巴系统在人类全生命周期健康和疾病中的重要性,识别其在大脑发育和衰老过程中的遗传因素至关重要。最近,一项全基因组关联研究(GWAS)确定了多个与通过磁共振成像(MRI)测量的血管周围空间(PVS)相关的遗传位点,而PVS是脑胶质淋巴系统的结构组成部分。除了PVS之外,弥散张量成像(DTI)分析沿血管周围空间(ALPS)指数是人类脑胶质淋巴系统相关状况的另一个非侵入性指标。与测量脑胶质淋巴系统形态的PVS不同,ALPS指数测量的是沿PVS方向的那些垂直投射纤维和联合纤维中液体的扩散性。与实验结果一致,越来越多的研究观察到在脑胶质淋巴功能受损的情况下,如在神经退行性疾病和阿尔茨海默病病理存在的情况下,ALPS指数降低,这表明ALPS指数在反映脑胶质淋巴功能方面具有潜力。评估脑胶质淋巴液体动力学。然而,据研究人员所知,ALPS指数的遗传机制尚未得到研究。
此外,脑胶质淋巴系统的发育是一个从胎儿期持续到成年期的长期过程。在啮齿动物中,脑胶质淋巴功能从新生期到成年期逐渐增加,但随着年龄的增长而下降。同样,在人脑中,ALPS指数随年龄增长而增加,在中年(40多岁)达到峰值,然后逐渐下降。因此,早期和晚期生命阶段ALPS指数变化的原因和后果可能不同,分别反映了大脑的发育和退化。研究人员假设与年轻大脑中ALPS指数相关的遗传因素可能与老年大脑中的遗传因素有显著差异。发现能够解释年轻和老化大脑中ALPS指数变化的遗传因素,可能为脑胶质淋巴系统的发育和老化过程提供新的见解,并有助于识别疾病风险和韧性。
在此,研究人员开展了全基因组关联研究(GWAS),以识别与英国生物样本库(UKB发现队列,N = 31,021,年龄45-82岁)中ALPS指数相关的遗传变异、位点和基因,并在两个独立样本(英国生物样本库非英国白人,N = 3,470,年龄45-81岁;ADNI,N = 260,年龄55-75岁)中验证了这些发现。由于此前观察到的在神经退行性疾病中双侧ALPS指数的差异是否由于测量偏差或具有临床意义尚不确定,研究人员分析了平均ALPS指数(主要表型),以及分别分析右侧和左侧半球的ALPS指数,作为相互验证或探索性分析。其次,研究人员利用英国生物样本库发现队列、ADNI以及包括ABCD(N = 4,307,年龄9-11岁)、IMAGEN(N = 1,589,年龄13-18岁)、HCP(N = 219,年龄22-36岁)在内的其他三个独立队列,探索了遗传效应对ALPS指数在全生命周期的一致性和差异性。进一步调查了与ALPS指数纵向变化相关的遗传因素。随后,研究人员进行了功能注释,以及与神经精神表型的遗传相关性和共享遗传决定因素的分析,以研究已识别位点的生物学意义。
2.结果
2.1 ALPS指数的提取与评估
本研究共纳入了来自五个队列(UKB、ADNI、IMAGEN、HCP和ABCD)的40,866名参与者,年龄范围从儿童到老年人(9-82岁,见表1)。发现GWAS分析包括来自31,021名英国生物样本库(UKB)脑成像样本的数据(53.0%为女性;年龄范围45-82岁;见表1),这些样本均为英国白人血统,且无流行的痴呆、中风、脑积水和颅内肿瘤。研究设计的示意图和指数处理过程的插图分别见补充图1和图1。ALPS指数是使用半自动化且高度可靠的流程从每个队列的弥散张量成像(DTI)中提取的。在ALPS指数提取中,对感兴趣区域(ROIs)放置的准确性进行了优先评估,这一操作在UKB和IMAGEN数据集中完成(方法)。研究表明,手动ROI和群体平均ROI模板所衍生的ALPS指数在老年人群(例如UKB参与者;皮尔逊相关系数r = 0.77,p = 1.45 × 10^-15)和年轻人群中(例如IMAGEN参与者;皮尔逊相关系数r = 0.81,p = 1.08 × 10^-12;见补充图2)具有很大的可比性。不同队列中ALPS指数的分布情况见表1和补充图3。研究人员先前的研究已基于UKB和ADNI参与者展示了生成的ALPS指数的合理性和临床相关性,其中ALPS指数与阿尔茨海默病风险、阿尔茨海默病生物标志物和认知功能相关。本研究进一步表明,较低的ALPS指数与年龄较大、男性、认知功能较差、白质高信号(WMH)较多、手握力较弱、体重指数较低和血压较高有关,但与教育、睡眠时间或惯用手无关(见补充数据1)。
表1 | 样本和表型描述
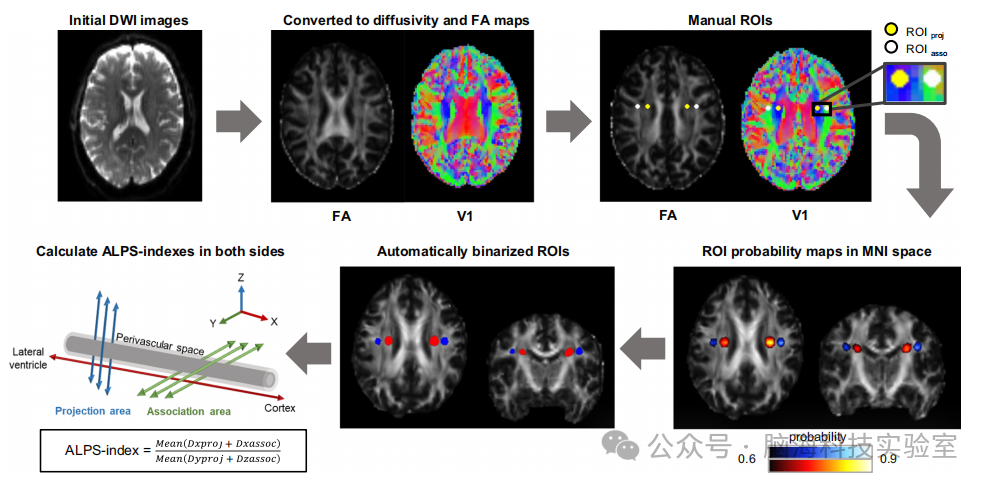
图1 | ALPS指数计算示意图
从初始弥散加权成像图像到沿血管周围空间指数计算的分析过程。选择脑室水平的横断面以显示与脑室体部垂直的投射纤维和联合纤维区域。彩色编码的各向异性分数(FA)图显示投射纤维(蓝色)和联合纤维(绿色)。在双侧投射纤维区域(黄色实心圆)和双侧联合纤维区域(白色实心圆)各放置了4个半径为4毫米的球形感兴趣区域(ROI)。然后,研究人员在标准空间中获得ROI概率图,并将其二值化为自动ROI。对于每个自动二值化的ROI,提取沿x轴、y轴和z轴的弥散性。ALPS指数计算为与纤维束垂直且与静脉平行的弥散性(Dxproj和Dxassoc)与与纤维束垂直且与静脉垂直的弥散性(Dyproj和Dzassoc)的比值。ALPS沿血管周围空间的分析,DWI弥散加权成像,FA各向异性分数,MNI蒙特利尔神经研究所,ROIasso联合纤维中的感兴趣区域,ROIproj投射纤维中的感兴趣区域。
2.2 ALPS指数的遗传发现
在对平均ALPS指数(mALPS-index)进行的全基因组关联分析中,涉及8,447,093个单核苷酸多态性(SNPs)的分位数-分位数(QQ)图显示了膨胀(连锁不平衡评分回归(LDSC):λGC = 1.159,平均χ² = 1.195;见补充图4和补充数据2)。然而,LDSC截距(1.005;标准误差(SE)= 0.023)和比例(2.7%)均表明,观察到的基因组膨胀是由于多基因结构,而不是由于人群分层造成的混杂。利用LDSC计算的基于单核苷酸多态性(SNP)的mALPS指数的遗传力(h²SNP)为28.4%(SE = 0.023),而通过GCTA计算为32.5%(SE = 0.014),这表明mALPS是一种可遗传的性状。
2.3 全基因组关联分析结果
对平均ALPS指数(mALPS-index)的全基因组关联分析在英国生物样本库(UKB)发现队列中识别出554个全基因组显著的单核苷酸多态性(SNPs,p < 5 × 10^-8,见补充数据3)。随后,研究人员利用FUMA流程发现了25个独立显著的变异(r² < 0.6)和15个主要变异(r² < 0.1),这些变异映射到15个独特的基因组位点(表2、图2A和补充数据4、5),范围从14.98到526.18千碱基对(图2B和补充图5)。所有主要变异均通过条件和联合(COJO)分析成功确认(见补充数据6)。接着,他们使用FINEMAP识别具有高后验因果概率(>0.6)的变异(见补充数据7)。NUAK家族SNF1样激酶1(NUAK1)的主要SNP rs12146713是所有位点中后验概率最高的变异(0.99),其次是位于SULF2基因中的SNP rs6012259(后验概率0.69)。候选SNPs的详细结果总结在图2C、补充结果和补充数据8-12中。为了探索罕见变异对ALPS指数的影响,研究者使用SKAT-O检验对UKB的全外显子组测序数据进行了罕见变异关联研究(RVAS)。基于基因的负担检验和单变异分析在经过Bonferroni校正后未发现任何显著关联,尽管观察到一些接近阈值的关联(见补充数据13、14)。其中最强的关联是基因HPS5与mALPS指数之间的关联(pSKAT-O = 9.74 × 10^-5)。
表2 | 与ALPS指数相关的遗传位点
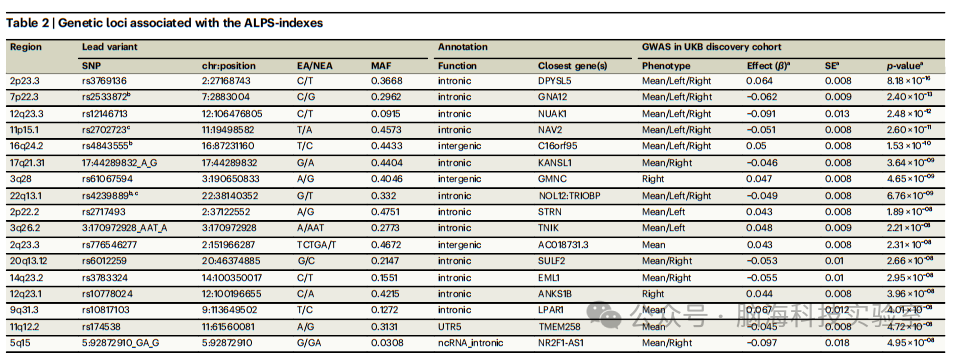
P值和β效应是通过PLINK在GWAS中使用线性加性回归模型(双侧检验)计算得出的,同时调整了年龄、年龄的平方、性别、成像中心和前十个遗传主成分(PCs)。显示的P值是原始值。GWAS分析2调整了年龄、年龄的平方、性别、成像地点和前十个遗传主成分。Chr染色体,EA效应等位基因,GWAS全基因组关联研究,NEA非效应等位基因,MAF次要等位基因频率,SE标准误差,SNP单核苷酸多态性。a这些值来自“表型”列中报告的第一个表型的GWAS结果。b对于这些位点,在右侧ALPS指数的分析中主要SNP不同(在平均ALPS指数的GWAS中 → 右侧ALPS指数的主要SNP:rs2533872 → rs2644291;rs4843555 → rs4843550;rs4239889 → rs1894526)。c对于这些位点,在左侧ALPS指数的分析中主要SNP不同(在平均ALPS指数的GWAS中 → 左侧ALPS指数的主要SNP:rs2702723 → rs2702721;rs4239889 → rs4820303)
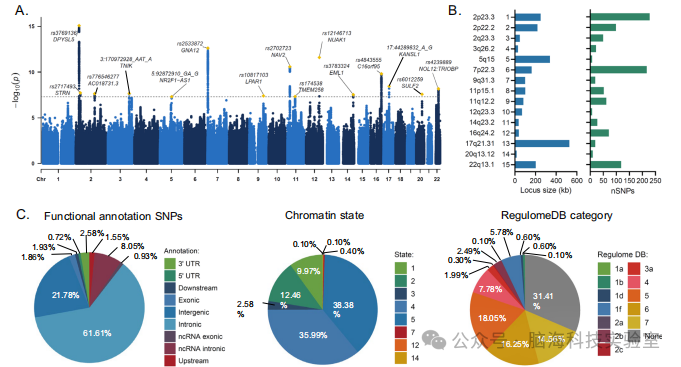
图2 | 基于SNP的与平均ALPS指数的关联及功能注释
英国生物样本库(UKB)发现队列中平均ALPS指数的全基因组关联研究(GWAS)结果的曼哈顿图。该图在y轴上显示每个SNP的 - log10转换P值,在x轴上显示沿染色体的碱基对位置(n = 37,327名个体)。显示的P值为双侧。虚线表示在进行线性加性回归模型的GWAS中达到全基因组显著性的P值(P < 5 × 10^-8)。显著位点以黄色正方形标记,并用其主要SNP进行标注。B 风险位点的大小和与平均ALPS指数相关的变异数量。C 饼图显示了与平均ALPS指数发现GWAS中全基因组显著主要SNP连锁不平衡(LD)的SNP的功能后果分布、127种组织和细胞类型中的最小染色质状态分布,以及RegulomeDB评分的分布(RegulomeDB评分是一个介于1a到7之间的分类评分,表示SNP作为调控元件的生物学证据,评分越低表示SNP作为调控元件的可能性越高)。SNP 单核苷酸多态性。
2.4 分层连锁不平衡评分回归(LDSC)分析
分层LDSC进一步显示,遗传力(h²)的显著富集源于背景选择(富集度 = 1.47,p = 5.21 × 10^-12)、CpG含量(富集度 = 1.15,p = 3.70 × 10^-5)、组蛋白标记(H3K27ac、H3K4me1和H3K9ac)以及位于保守区域(GERP NS;富集度 = 1.86,p = 8.01 × 10^-6)和超级增强子(富集度 = 2.29,p = 5.55 × 10^-6)的SNPs(见补充数据15),这表明存在表观遗传基因调控。
由于mALPS指数是左右半球ALPS指数的平均值,研究人员进一步通过比较mALPS指数与左右ALPS指数的遗传信号来检验ALPS指数的不对称性。在左右ALPS指数的分析中,结果与mALPS指数的结果高度相关(见补充数据2-5)。在右侧ALPS指数的分析中,额外发现了两个风险位点:3q28(GMNC)和12q23.1(ANKS1B)(见表2、补充图4-6和补充数据4)。三种ALPS指数的表型和遗传相关性均显著(见补充数据16)。为了验证遗传关联分析中独立显著SNPs的稳健性,他们进行了敏感性分析。如补充数据17所示,当将平均各向异性分数(FA)和平均弥散率(MD)作为额外协变量时,85.7%的SNPs仍保持全基因组显著,这表明白质完整性对结果的潜在混杂影响极小。
2.5 ALPS指数在全生命周期的遗传结构
在两个独立样本中进行了验证:3,470名英国生物样本库(UKB)的非英国白人参与者,他们未被纳入发现队列,以及260名来自阿尔茨海默病神经影像学倡议(ADNI)队列的非西班牙裔白人非痴呆参与者(见表1和补充图7)。在发现队列中识别的25个独立显著风险变异中,有14个(60.0%)变异在至少一个验证队列中得到确认(p < 0.05)。其中8个变异在经过假发现率(FDR)校正后达到显著性,且其效应方向与发现分析中观察到的一致(见补充数据5)
2.6 ALPS指数在全生命周期的遗传结构
由于ALPS指数与年龄显著相关(见补充图8、9和补充数据18),研究者假设某些遗传位点可能在全生命周期对ALPS指数有不同的影响。为了验证这一假设,他们在年轻成年人的独立队列中对已识别的独立显著变异与ALPS指数之间的关联进行了检验,作为泛化分析。研究涉及三个泛化队列:ABCD(N = 4,307,平均年龄9.91 ± 0.62岁)、IMAGEN(N = 1,589,13.97 ± 0.40岁)和HCP(N = 219,28.89 ± 3.68岁)(见表1)。在至少一个队列中,与发现结果效应方向一致的mALPS指数的7个风险变异显示出名义上的显著关联,其中4个在经过假发现率(FDR)校正后达到显著性(见补充数据5)。这些一致的发现表明,遗传关联在整个生命周期内是稳健的。例如,2p23.3位点的变异在不同年龄队列中与mALPS指数一致相关,除了样本量最小的HCP和ADNI(见图3A)。从英国生物样本库(UKB)发现GWAS变异中衍生的多基因风险评分(PRS)与mALPS指数显著相关(在英国生物样本库非英国白人亚组中β = 0.19,p = 1.89 × 10^-31;在ADNI中β = 0.14,p = 0.02),但在泛化队列中则不然。
2.7 独立显著变异的遗传效应
随后,在来自四个独立队列的七个亚组中测试了独立显著变异的遗传效应:ABCD(年龄范围9-11岁)、IMAGEN(13-18岁)、HCP(22-36岁)、ADNI(55-75岁),以及三个英国生物样本库(UKB)亚组(按样本量三分位数划分的年龄范围:45-61岁、62-68岁、69-82岁;见补充数据19)。在对等位基因进行校准后,使用线性模型和二次模型对遗传效应按年龄进行元回归分析。分析识别出六个变异在经过假发现率(FDR)校正后显示出与年龄的线性或二次关联(见补充数据20)。值得注意的是,mALPS指数中20q13.12位点的主要变异rs6012259可能参与神经退行性过程,因为其负效应在老年人群中更为显著(见图3B)。16q24.2位点变异的遗传效应与年龄呈二次关联(见图3C)。
利用来自年轻队列(ABCD:N = 2,152,基线年龄9.91 ± 0.61岁)和老年队列(UKB:N = 3,580,基线年龄62.07 ± 7.35岁)的纵向弥散张量成像(DTI)数据,研究人员观察到在ABCD队列中,mALPS指数在平均(标准差)随访1.99(0.14)年后增加(配对Wilcoxon检验p = 6.7 × 10^-4),而在UKB队列中,mALPS指数在平均(标准差)随访2.60(0.99)年后减少(配对Wilcoxon检验p < 2.22 × 10^-16)(见补充图10)。通过ALPS指数测量的胶质淋巴功能在年轻大脑中随时间积累(mALPS指数年变化率:0.004 ± 0.06),但在老年大脑中随时间下降(-0.005 ± 0.02)(见表1)
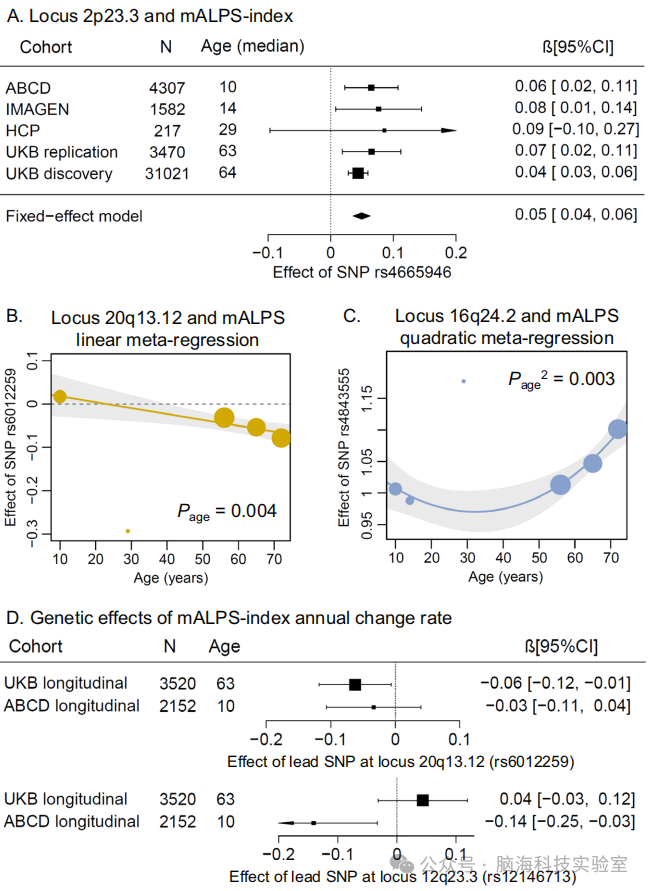
图3 | 全生命周期内ALPS指数的遗传效应及其年变化率
A 2p23.3位点rs4665946(2:27347931)对平均ALPS指数的固定效应元分析。B 和 C 使用线性模型和二次模型对SNP效应按年龄进行元回归分析。显示的P值为双侧,未进行多重比较校正。图中显示了通过假发现率(FDR)校正的两个变异的结果。x轴表示每个队列的中位年龄,y轴表示对平均ALPS指数的遗传效应。点的大小表示对平均ALPS指数的遗传效应的 - log10 p值。回归线周围的误差带表示95%置信区间。D 森林图显示了20q13.12位点rs6012259(20:46374885)和12q23.3位点rs12146713(12:106476805)的遗传效应。CI 置信区间,SNP 单核苷酸多态性,mALPS 平均ALPS指数。
2.8 独立显著变异的纵向验证
研究人员检验了在发现GWAS中识别的与mALPS指数相关的独立显著变异是否能在其年变化率分析中得到验证。在ABCD队列中,两个变异(rs4757810和rs12146713)和在UKB队列中的一个变异(rs6012259)显著(P < 0.05),且与发现GWAS中的效应方向一致(见图3D和补充数据21)。在纵向分析和按年龄进行的元回归分析中,20q13.12位点的主要SNP(rs6012259)在神经退行性过程中对mALPS指数的影响显著强于在神经发育过程中的影响(见图3B、D和补充数据20、21)
2.9 与神经精神表型和血管危险因素的遗传关联
研究人员通过在NHGRI-EBI GWAS目录和先前发表的PVS研究中进行关联查询,检验了与ALPS指数相关的变异是否与其他表型相关。在与ALPS指数相关的17个位点中,有四个(12q23.1、14q23.2、2q23.3和3q26.2)此前未被报道,而其余13个位点此前均已被报道与认知测量(2p22.2、2p23.3、5q15、17q21.31和22q13.1)、神经精神特征(2p23.3、3q28、11q12.2和17q21.31)以及心血管特征(2q22.2、2p23.3、11q12.2、14q32.2和17q21.31)共享遗传影响的结构遗传位点有关(见补充数据22)。特别是,在mALPS GWAS中,9q31.3位点(LPAR1)的主要变异(rs10817103)显示出与WM-PVS负荷相关的全基因组显著p值(P = 3.7 × 10^-13)(见补充数据23)
2.10 ALPS指数与脑结构、白质完整性和脑功能的遗传相关性
研究者利用连锁不平衡分数回归(LD-score regression)更好地理解ALPS指数与脑结构、白质完整性和脑功能之间的相关性(见补充数据24)。所有脑室体积测量指标均与三个ALPS指数显著相关(见图4A),其中与两侧脑室体积(rg = -0.47至-0.42,PBonferroni = 4.67 × 10^-19至3.64 × 10^-16)的相关性最为显著。关于脑微结构(见图4B),与三个ALPS指数相关性最显著的是穹窿(rg = 0.42-0.43,PBonferroni = 1.42 × 10^-14至2.92 × 10^-12)。然而,在经过Bonferroni校正后,未发现与脑功能特征有显著相关的结果(见补充数据24)。
随后,研究人员检验了ALPS指数与其他神经精神表型和血管危险因素之间的遗传相关性,包括潜在危险因素、认知功能、神经精神和神经系统疾病、阿尔茨海默病的脑脊液生物标志物以及脑小血管病(cSVD)的MRI标志物(见补充数据25)。尽管在应用Bonferroni校正后未观察到统计学显著的关联,但在名义上观察到白质高信号(WMH)体积(rg = -0.27至-0.26,P = 0.008-0.012)与所有三个ALPS指数之间存在显著相关性。左手握力更大、反应时间更短以及脉压更高在名义上与至少一个ALPS指数存在遗传相关性(见图4C)
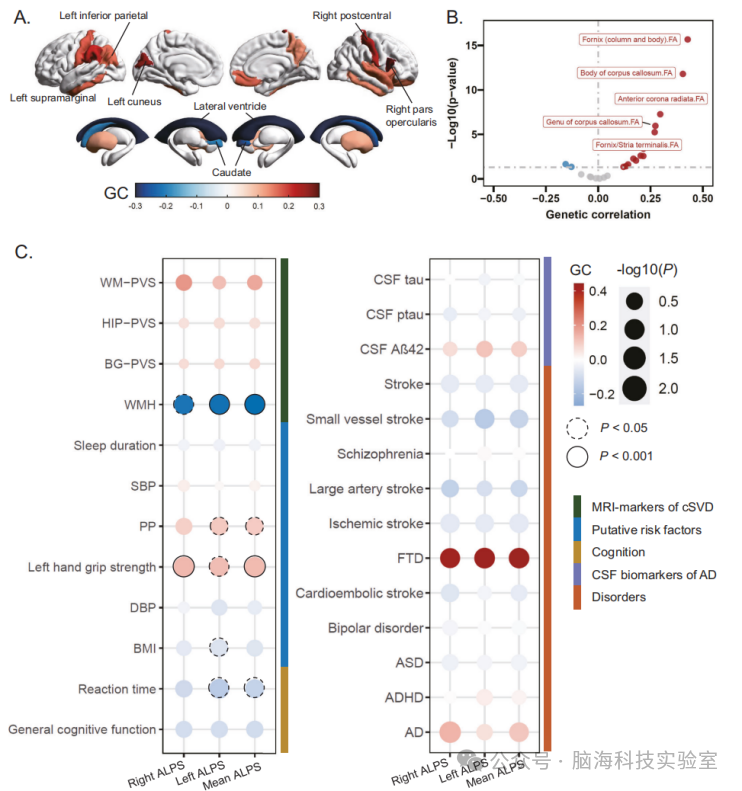
图4 | 识别的ALPS指数位点的遗传相关性
A 平均ALPS指数与脑体积之间的遗传相关性。所有与平均ALPS指数在名义P < 0.05水平上具有遗传相关的区域均根据正(红色)或负(蓝色)遗传相关性估计值的大小进行着色。通过假发现率校正的遗传相关性估计值的区域用文字标注。
B 火山图显示了平均ALPS指数与白质微结构之间的遗传相关性估计值(x轴)和 - log10(p值)(y轴)。图中标签显示了五个最显著的白质微结构。
C 气泡图显示了三个ALPS指数与潜在危险因素、认知测量和其他大脑衰老的MRI标志物之间的遗传相关性。点的颜色表示正(红色)或负(蓝色)遗传相关性的大小,而点的大小表示遗传相关性的 - log10(P)。具有名义P < 0.05的遗传相关性估计值用虚线圆圈标注,而通过假发现率校正的估计值用实线圆圈标注。遗传相关性是使用LDSC估计的。所有显示的P值均为双侧,且为未经多重比较校正的原始P值。
AD 阿尔茨海默病,ADHD 注意力缺陷多动障碍,ASD 自闭症谱系障碍,BG-PVS 基底神经节血管周围空间,BP 双相情感障碍,cSVD 脑小血管病,FTD 额颞叶痴呆,BMI 体重指数,CSF 脑脊液,DBP 舒张压,FA 各向异性分数,FDR 假发现率,GC 遗传相关性,HIP-PVS 海马血管周围空间,MRI 磁共振成像,SBP 收缩压,PP 脉压,WM-PVS 白质血管周围空间,WMH 白质高信号。
2.11 ALPS指数的多基因结构及与神经精神表型和血管危险因素的遗传重叠
随后,研究人员利用条件假发现率(condFDR)和联合假发现率(conjFDR)分析,进一步探索了ALPS指数的多基因结构及其与神经精神表型和血管危险因素的潜在遗传重叠。在condFDR < 0.01的条件下,基于与神经精神表型和血管危险因素的关联显著性,ALPS指数的位点发现能力显著增强,识别出的遗传位点数量超过了发现GWAS的结果(图5A中每个 >15)。条件Q-Q图显示ALPS指数与精神分裂症(SCZ)、脑室周围血管周围空间(HIP-PVS)、白质高信号(WMH)、脑脊液pTau和脑脊液tau之间存在明显的遗传重叠(图5B和补充图11)。在conjFDR < 0.05的阈值下,识别出ALPS指数与主要精神障碍、脑脊液(p)Tau和脑小血管病(cSVD)生物标志物之间的几个共享位点(图5A、补充数据27和补充图12)。
2.12 双向Mendelian randomization分析
研究人员进一步进行了双向Mendelian randomization(MR)分析,以检验潜在的因果关系。在正向MR中,左侧ALPS指数水平的增加与经过Bonferroni校正后的注意力缺陷多动障碍(ADHD)风险降低相关(OR [95% CI] = 0.647 [0.512–0.818],PIVW-Bonferroni = 0.009)(补充数据28)。在反向MR中未发现显著结果(补充数据29)
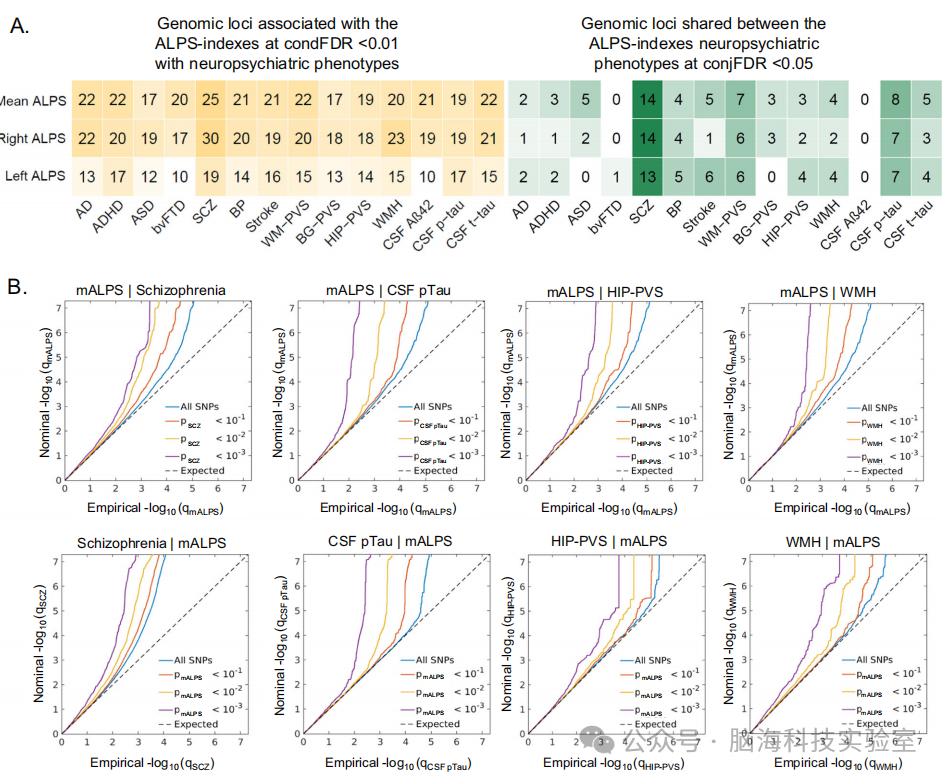
图5 | ALPS指数与神经精神和神经系统疾病及生物标志物的多基因重叠
A 通过条件假发现率(condFDR)分析,在神经精神和神经系统疾病及生物标志物的条件下,增强了对三个ALPS指数的遗传位点的发现。联合假发现率(conjFDR)分析检测了三个ALPS指数与神经精神疾病和生物标志物之间的共享遗传位点。
B 平均ALPS指数在精神分裂症、脑脊液pTau、海马血管周围空间(HIP-PVS)、白质高信号(WMH)等条件下的条件Q-Q图,展示了遗传重叠。在标准GWAS阈值p < 5 × 10^-8以下的表型中,名义与经验 - log10 p值(校正膨胀)的条件Q-Q图,作为与另一个表型关联显著性的函数,分别在p ⩽ 0.1、p ⩽ 0.01、p ⩽ 0.001水平上。蓝色线表示所有SNPs。虚线表示零假设。
AD 阿尔茨海默病,ADHD 注意力缺陷多动障碍,ASD 自闭症谱系障碍,BG-PVS 基底神经节血管周围空间,BP 双相情感障碍,bvFTD 行为变异型额颞叶痴呆,CSF 脑脊液,HIP-PVS 海马血管周围空间,mALPS 平均ALPS指数,SCZ 精神分裂症,WM-PVS 白质血管周围空间,WMH 白质高信号。
2.13 基因水平分析与功能富集
为了深入了解与ALPS指数相关的基因,研究者利用全基因组基因基础关联研究(GWGAS,采用MAGMA;见图6A)以及通过FUMA进行的物理位置、表达数量性状位点(eQTL)和3D染色质相互作用映射策略,开展了基因水平分析。基于英国生物样本库(UKB)发现队列中三个ALPS指数的GWAS结果,这三种映射策略共识别出151个独特基因(见补充数据30)。GWGAS在全基因组显著水平(p < 2.65 × 10^-6)上识别出40个独特基因,其中30个与FUMA所涉及的基因重叠,从而总共确定了161个与ALPS指数相关的独特基因(见图6B和补充数据31)。其中,111个(68.9%)基因在三个ALPS指数之间重叠(见图6C),并且有21个基因具有最有力的证据,因为它们同时被所有四种方法所涉及(见补充数据32)。
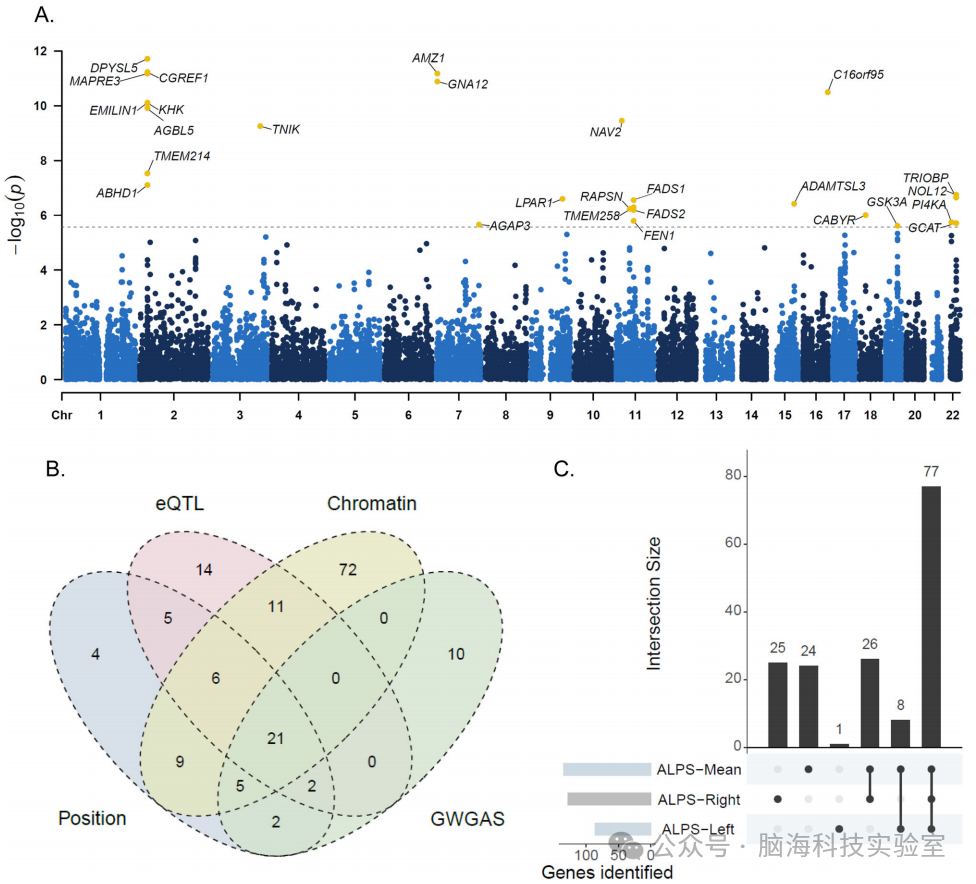
图6 | 通过GWGAS和基因映射策略识别的与ALPS指数相关的基因
A 平均ALPS指数的全基因组基因基础关联研究(GWGAS)的曼哈顿图(n = 37,327名个体)。P值通过MAGMA基因基础检验计算得出。y轴显示每个基因的 - log10 p值。x轴显示染色体位置(起始位置)。虚线表示基因基础检验的全基因组显著性的Bonferroni校正阈值(P < 2.65 × 10^-6;0.05/18,878)。
B 韦恩图显示了通过位置映射、eQTL映射、染色质相互作用映射和GWGAS识别的与三个ALPS指数(平均、左侧和右侧)相关的基因数量。
C Upset图显示了三个ALPS指数(平均、左侧和右侧)之间的独特和共有基因数量。
eQTL 表达数量性状位点,GWGAS 全基因组基因基础关联分析。
2.14 细胞类型富集分析与功能分析
为了评估基因是否在特定脑细胞类型中过度表达,研究人员进行了细胞类型富集分析。对于平均ALPS指数(mALPS-index),显著信号在少突胶质细胞祖细胞(OPC)和少突胶质细胞中富集(见补充数据33)。少突胶质细胞是非神经元细胞,它们为轴突提供支持,并在运动技能学习和记忆形成中发挥作用。
为了探究与ALPS指数相关的所有已识别基因的生物学特性,研究者利用g:Profiler网络工具中的g:GOSt功能进行了富集分析。功能分析显示,在神经发生、解剖结构发育和胶质细胞发育的生物学过程中富集,以及在细胞连接和细胞突起的细胞组分中富集,还在氧化还原酶活性和亚油酰辅酶A去饱和酶活性的分子功能中富集(见补充数据34)。FUMA富集分析揭示了与神经精神表型和代谢相关的几种富集表型,包括帕金森病、认知能力、神经质和脂质代谢(见补充数据35)。此外,他们在GWGAS定义的基因内进行了功能分析,随后进行了MCL聚类。发现一个基因簇(ARHGAP27、ARL17B、CRHR1、KANSL1、PLEKHM1、SPPL2C、STH)在神经退行性、认知功能和神经精神障碍中富集(见补充数据36)。
2.15 基因 - 药物相互作用分析
为了探究这些已识别基因是否可能成为药物治疗的潜在靶点,研究人员随后利用药物基因相互作用数据库(DGIdb)搜索了基因-药物相互作用。这一搜索识别出25个独特基因,涉及781种独特药物(见补充数据37)。其中,MAPT基因报告的相互作用数量最多(445种相互作用),其次是FEN1基因(199种相互作用)。观察到几个潜在的药物治疗干预靶点(见补充数据38),涵盖82个独特基因。其中,60个(73.2%)此前未被报道与已知药物相互作用。
2.16 基因表达模式分析
研究人员研究了与ALPS指数相关的基因从发育到成年的脑表达模式。一些基因在整个生命周期内的表达水平存在显著变化,某些基因在胎儿期达到峰值(例如NAV2、TRIOBP、FEN1、MAPT、DPYSL5)(见补充图13),这表明遗传决定的对胶质淋巴活动的影响可能始于胎儿期。
3.讨论
本研究调查了多达40,866名9至82岁参与者的ALPS指数的遗传结构。ALPS指数显示出中等程度的遗传性。研究人员发现了17个独特的全基因组显著风险位点与ALPS指数相关,主要与平均ALPS指数(mALPS-index)相关,包括右侧ALPS指数的两个位点。通过基因水平分析,共识别出161个独特的候选基因。与研究者的假设一致,一些已识别的遗传信号在大脑发育和衰老过程中显示出变化效应。功能分析显示,在神经发生、解剖结构发育和胶质细胞发育的生物学过程中富集,以及在神经精神疾病和代谢的表型中富集。观察到ALPS指数与更好的认知表现、更好的身体指标、较小的脑室体积和较低的脑小血管病(cSVD)MRI标志物之间存在遗传相关性。ALPS指数与神经精神和神经系统疾病、cSVD的MRI标志物以及阿尔茨海默病的脑脊液生物标志物之间存在遗传重叠,表明这些表型之间存在遗传多效性,可能共享共同的胶质淋巴病理机制。Mendelian randomization分析进一步强调,较高的ALPS指数与注意力缺陷多动障碍(ADHD)风险降低相关。这些结果增强了对胶质淋巴清除活动的神经生物学理解,并为功能实验提供了具体线索。
3.1 12q23.3位点的强证据
研究人员发现了12q23.3位点的强证据,该位点由rs12146713(NUAK1)标记,其因果概率最高(0.99),这是通过FINEMAP检测到的。NUAK1基因是AMP激活蛋白激酶家族丝氨酸/苏氨酸激酶的成员,已被发现与穹窿白质微结构相关。穹窿在空间记忆、情景记忆和执行功能的形成中起作用。这与本研究发现的ALPS指数与穹窿白质微结构之间的遗传相关性以及先前观察到的ALPS指数与认知之间的关联一致。此外,NUAK1蛋白通过选择性磷酸化tau蛋白的Ser356位点来调节tau水平,而p-tau Ser356与阿尔茨海默病病理进展有关。抑制NUAK1可以防止tau蛋白积累和神经纤维缠结的形成,并在tau病小鼠模型中挽救记忆缺陷。值得注意的是,NUAK1基因在大脑中的表达随年龄增长而增加,在20多岁时达到峰值。研究发现,rs12146713-C(NUAK1)等位基因与英国生物样本库(UKB)中较低的mALPS指数以及ABCD中mALPS指数增长较慢有关。这表明该基因对老年大脑中胶质淋巴活动的降低和年轻大脑中胶质淋巴功能的缓慢发展具有遗传效应。因此,NUAK1基因在早期生活中的相对低表达与胶质淋巴发育过程一致,而其随年龄增长的表达增加可能与随后的年龄依赖性胶质淋巴活动下降有关。NUAK1基因是可成药的,尽管目前尚无针对该基因的药物治疗用于神经或精神疾病的管理。这些发现共同指向NUAK1作为神经精神疾病的潜在治疗靶点,调节胶质淋巴清除活动可能是其额外的作用机制。
3.2 ALPS指数的纵向变化与年龄依赖性遗传效应
利用来自多个队列的纵向数据,研究人员观察到儿童大脑中的mALPS指数随时间增加,而老年大脑中的mALPS指数随时间减少。先前的研究强调了与年龄相关的脑胶质淋巴活动的变化以及年龄依赖性遗传效应对人脑神经影像特征的重要性。鉴于通过ALPS指数量化的脑胶质淋巴活动具有年龄依赖性,研究人员在老年队列中识别的遗传关联主要代表了胶质淋巴系统的退化过程,而在年轻队列中可能主要涉及成熟过程。正如研究者假设的那样,年轻和老年大脑中ALPS指数的遗传驱动因素可能不同,因此在英国生物样本库(UKB)发现队列中识别的大多数独立遗传信号在年轻队列中并不显著。从发现GWAS汇总数据中衍生的多基因风险评分(PRS)在年轻队列中未显示出显著性。这不太可能是由于这些年轻队列样本量小,因为在另一个样本量较小的老年个体队列(ADNI)中构建的PRS得出了显著结果。尽管有这些发现,但2p23.3和3q26.2位点的几个遗传信号在年轻和老年个体中对胶质淋巴活动均显示出显著影响。在按年龄进行的元回归分析中,这些变异未显示出不一致的效应,这进一步表明它们在整个生命周期内具有一致的作用。
3.3年龄依赖性关联与基因功能
本研究通过按年龄进行的元回归分析和纵向GWAS,发现几个位点与mALPS指数存在显著的年龄依赖性关联。20q13.12位点的主要变异rs6012259(SULF2)的G等位基因与年轻成年人和老年人群(即英国生物样本库(UKB)和人类连接组计划(HCP)队列)中的较低mALPS指数相关,且在老年时负效应更强。在包含中年至老年人的UKB纵向队列中,该变异还与mALPS指数更快的下降相关。在ABCD队列的儿童中,20q13.12位点的显著性减弱。这些发现表明,该位点可能主要影响老年人的胶质淋巴功能障碍。此外,SULF2基因编码一种参与多种肝素硫酸蛋白多糖依赖的受体酪氨酸激酶信号通路的酶。它在中枢神经系统中高表达,并在调节神经突起生长和大脑稳态中发挥作用。此外,20q13.12位点的变异rs4757810(NAV2)的A等位基因与UKB中的较低mALPS指数以及ABCD纵向队列中mALPS指数增长较慢相关。它可能在早期生活中对胶质淋巴系统的发育产生不利影响,并在晚年加剧胶质淋巴功能障碍。NAV2基因在神经系统发育和血压调节中起关键作用。此前,它被建议为阿尔茨海默病(AD)的风险基因,与AD中的情景记忆表现以及AD的发病年龄和风险相关。此外,研究揭示了在儿童和老年人群中识别的遗传位点效应存在差异。例如,9q31.3位点(由LPAR1的rs10817103标记)仅在中年和老年队列(即ADNI和UKB)中与mALPS指数存在显著关联,但在任何年轻队列中均未发现。一种推测是表观遗传调控,随着年龄增长,表观遗传标记(如DNA甲基化和组蛋白修饰)的变化可能导致某些位点的表达发生改变,从而影响晚年的表型。这些位点的表观遗传调控可能在未来成为针对人类疾病胶质淋巴靶向治疗的靶点。
3.4 ALPS指数的遗传结构与左右半球差异
左右两侧ALPS指数以及平均ALPS指数(mALPS-index)的遗传结构高度相似。三者之间存在显著的遗传和表型相关性,且在遗传力和已识别的遗传位点方面结果高度一致。然而,较高的左侧ALPS指数与更好的认知功能、较低的血压和体重指数(BMI)存在显著的遗传关联,而右侧ALPS指数则未观察到这种关联。先前的队列研究已表明,神经退行性疾病患者左侧半球的ALPS指数下降更为显著,但双侧ALPS指数之间并无显著差异。尽管先前的证据支持双侧ALPS指数在临床相关性上存在差异,但不能排除测量偏差可能是这些差异的原因。惯用手被认为是潜在的影响因素。然而,在具有不同惯用手的参与者之间,双侧ALPS指数并无显著差异。从遗传学角度来看,鉴于双侧与平均ALPS指数的三种遗传结果之间存在细微差别,仍有必要分别对这三种特征进行深入研究。进一步的实验研究也有助于阐明这些差异是否具有病理生理学意义,还是仅仅是测量误差的结果。
3.5 胶质淋巴清除活动的遗传决定因素
本研究利用基于弥散张量成像(DTI)的非侵入性ALPS指数来探索胶质淋巴清除活动的遗传决定因素。人类胶质淋巴系统的另一种非侵入性测量方法是血管周围空间(PVS),这是在磁共振成像(MRI)上可见的胶质淋巴系统的一部分。ALPS指数和PVS之间存在多个重叠的位点,特别是LPAR1(9q31.3)。该位点与mALPS指数和白质PVS均显示出全基因组显著关联,并被认为是中枢神经系统疾病的潜在治疗靶点。例如,由LPAR1编码的环状RNA可能通过吸附其靶向微小RNA来调节神经元细胞死亡和神经炎症,从而增加阿尔茨海默病(AD)的风险。然而,在白质、海马或基底神经节中,ALPS指数与PVS负荷之间均未观察到显著的遗传相关性。尽管PVS的异常扩张和ALPS指数的降低都被假设与胶质淋巴功能障碍有关,但ALPS指数的临床相关性与PVS负荷并不相当。ALPS指数的异常与AD中的病理蛋白积累和认知衰退密切相关,甚至先于并预测加速的Aβ沉积。而PVS负荷与脑小血管病的状况更为密切相关,但与AD病理机制无关。虽然这超出了本文的范围,但一个可能的解释是,ALPS指数和PVS负荷的变化可能由胶质淋巴活动之外的其他不同的病理生理过程所支撑。
3.6 ALPS指数、脑脊液tau病理和脑室体积的共同遗传机制
结果表明,ALPS指数、脑脊液(CSF)tau病理和脑室体积之间存在共同的潜在遗传机制。ALPS指数测量沿血管周围空间(PVS)的液体扩散性,这可能代表胶质淋巴系统的功能活动。与相应的生理活动一致,遗传相关性分析揭示了较小的ALPS指数与较大的脑室体积之间存在强关联,这可能表明脑脊液流动受阻和神经退行性变化。此外,观察到ALPS指数与神经退行性疾病两个关键生物标志物——脑脊液pTau和Tau之间存在遗传重叠。胶质淋巴系统在清除脑实质中的tau蛋白种类方面发挥重要作用。联合假发现率(conjFDR)分析识别出几个重叠位点,包括两个显著的基因组位置:GMNC(3q28)和C16orf95(16q24.2)。这两个基因此前已被强调与脑脊液pTau水平和脑室体积相关,这意味着相同的遗传风险因素对这两种表型以及ALPS指数均有贡献,进一步强化了胶质淋巴活动与tau病理和神经退行性变化相关的观点。对于这两个位点,在本研究中与ALPS指数增加相关的等位基因也与较小的脑室体积但较高的脑脊液(p)Tau水平相关。此前曾讨论过这两个位点的反直觉效应。一个可能的解释是补偿假说。与tau病理相关的遗传因素导致脑脊液(p)Tau水平升高,这反过来又诱导液体清除过程和脑室缩小的代偿性增加,以清除废物并维持大脑储备。
3.7 ALPS指数与脑结构及功能特征的遗传相关性
遗传相关性分析表明ALPS指数与脑结构和功能特征密切相关。具体而言,ALPS指数与穹窿白质和反应时间显著相关,这是一对显著相关的表型。一个有趣的发现是较低的ALPS指数与较高的血压显著相关。血压可能会影响大脑的血流动力学,进而影响沿血管周围空间(PVS)的水分子运动,这正是ALPS指数所指示的。尽管已知ALPS指数与血管危险因素存在关联,但目前尚无证据表明ALPS指数与血压之间的关系。需要进一步的动物研究来验证这一发现,并探索其背后的机制。另一个与ALPS指数显著相关的表型是手握力,它与认知能力和痴呆风险有关。此外,还发现了ALPS指数与精神分裂症(SCZ)之间存在多基因重叠。研究表明,在成年期,精神分裂症患者的脑沟和蛛网膜下腔脑脊液水平增加。在进一步研究大脑液体运输活动在SCZ中的作用之前,ALPS指数与SCZ之间的多基因重叠为它们的共同基础提供了见解。
3.8 ALPS指数与ADHD风险的因果关系
一个值得注意的发现是ALPS指数与ADHD风险之间的单向因果关系,表明较高的ALPS指数可能对ADHD具有潜在的保护作用。影像学证据表明,ADHD儿童存在胶质淋巴功能障碍。Mendelian randomization(MR)分析结果支持胶质淋巴系统在ADHD发病机制中的保护作用。ADHD患者中改变的胶质淋巴活动的可能病理生理机制可能与神经炎症有关。胶质淋巴网络与下游的脑膜淋巴系统一起,作为大脑与外周免疫之间的交叉对话途径。受损的大脑清除途径可能导致从血液中招募免疫细胞和从大脑中清除大分子的过程出现异常。尽管有这些推测,但仍需要进一步探索以充分理解神经发育疾病中改变的胶质淋巴系统的病理生理学。
3.9 研究局限性
研究人员承认存在若干局限性。首先,尚未完全验证ALPS指数是否反映了人类的胶质淋巴清除活动。尽管先前的研究已表明ALPS指数具有高度的可重复性,并且与通过鞘内造影剂给药评估的胶质淋巴功能强相关,但尚未在病理生理学研究中得到彻底验证。在解释与ALPS指数相关的已识别遗传位点与胶质淋巴功能之间的关系时需谨慎。未来的研究方向将是整合多种胶质淋巴系统的方法,以及进行全转录组关联研究,以进一步阐明胶质淋巴功能的遗传基础。其次,研究队列未涵盖胎儿期和婴儿期的个体,以及青年和中年期的个体,表明整个生命周期的覆盖不够全面。第三,各队列之间的年龄重叠较少,样本收集、成像和基因分型方法存在差异。因此,在元回归分析中观察到的差异可能受到年龄以外的残留混杂因素的影响。第四,尽管研究者能够在两个独立队列——英国生物样本库(UKB)非英国白人和ADNI队列中验证多基因风险评分(PRS)和一部分遗传位点,但某些遗传变异未能显示出足够的可复制性。尽管这可能归因于这些位点的稳健性不足,但由于样本量小导致的统计功效有限而产生的样本异质性不容忽视。大规模样本将有助于验证我们的发现并取得新的发现。第五,UMA流程中使用默认参数来识别独立显著变异和候选变异,这采用了相对宽松的r²阈值。
4.总结
本研究识别了17个独特的与ALPS指数相关的遗传风险位点和161个基因,ALPS指数是胶质淋巴清除活动的一个有前景的标志物。研究人员检验了全生命周期内与年龄相关和与年龄无关的遗传效应,并展示了与神经精神表型的遗传关联。本研究的发现加深了对ALPS指数遗传结构的理解,并为进一步研究胶质淋巴清除活动在全生命周期内的生物学基础及其与大脑健康的相关性提供了思路。
5.方法
5.1 研究被试
本研究利用了来自多个队列参与者的原始影像和遗传数据:英国生物样本库(UKB,年龄范围45-82岁)、阿尔茨海默病神经影像学倡议(ADNI,年龄范围55-75岁)、青少年大脑认知发展研究(ABCD,年龄范围9-11岁)、青少年心理健康研究(IMAGEN,年龄范围13-18岁)和人类连接组计划(HCP,年龄范围22-36岁)。在数据收集之前,所有参与者均获得了知情同意,并且每个研究地点均获得了其各自的当地伦理委员会或机构审查委员会的批准。
从约40,000名中年至老年英国生物样本库(UKB)参与者中获取了原始影像数据,申请编号为19542。英国生物样本库是一项大型、基于人群的前瞻性队列研究,旨在调查影响中年和老年人疾病的遗传和非遗传因素。自2014年以来,已收集了多模态神经影像数据。主要分析集中在英国白人血统的英国生物样本库个体(如“ukb_sqc_v2.txt”中定义),并使用被认定为白人但非英国血统的英国生物样本库个体的独立子集进行内部验证。ADNI队列也由中年至老年参与者组成,用于外部验证。为了将主要结果推广到年轻群体,研究人员还利用了儿童(ABCD队列)、青少年(IMAGEN队列)和年轻成年人(HCP队列)的数据。为了尽量减少人群分层的影响,所有队列的参与者均被限制为白人血统。所有参与者的详细特征见表1。
5.2 基因分型与质量控制
英国生物样本库(UKB)团队集中进行了单核苷酸多态性(SNP)阵列基因分型、定相、插补和质量控制(QC)。研究人员使用PLINK v2.0进行了额外的质量控制程序,包括移除基因分型呼叫率低于0.95、自我报告性别与遗传性别不一致、存在性染色体异常、至少有十个三代以内亲属或杂合率异常的个体。在变异水平上,研究者排除了呼叫率低于0.95、次要等位基因频率(MAF)小于0.01、哈迪-温伯格平衡(HWE)P值小于10^-6或插补质量评分低于0.5的变异,最终得到850万个常染色体变异。对于ADNI、ABCD、IMAGEN和HCP,研究人员获得了原始基因型数据,并进行了与UKB类似的定相、插补和质量控制程序。基于英国生物样本库的全外显子组测序(WES)数据进行了罕见变异关联研究(RVAS)。不同队列的GWAS基因分型平台、插补面板和基因型质量控制的详细信息见补充方法和补充数据39。
5.3 弥散张量成像(DTI)和ALPS指数计算
在英国生物样本库(UKB)发现队列中,影像采集、预处理和质量控制由UKB代表开发并运行(https://biobank.ctsu.ox.ac.uk/crystal/crystal/docs/brain_mri.pdf)。简而言之,弥散MRI采用标准(“单极”)Stejskal–Tanner脉冲序列(回波时间 = 92毫秒),包含36个切片(分辨率 = 2.0 × 2.0 × 2.0毫米;视野 = 104 × 104 × 72矩阵),在50个不同的弥散加权方向上进行扫描(b值 = 1000 s/mm²)。预处理质量控制包括对模态进行对齐和去除伪影。所有扫描均进行了涡流校正和头部运动校正。随后应用了梯度失真校正。复制和泛化队列的影像细节见补充方法。在UKB和ABCD队列中,基线和随访的MRI协议相同。
Taoka等人开发并验证的半自动化且高度可靠的流程被用于从每个队列的弥散张量成像(DTI)中计算ALPS指数。在英国生物样本库(UKB)中,使用DTIFIT(版本2.0;FSL弥散张量拟合程序,FMRIB)处理沿x轴(右-左)、y轴(前-后)、z轴(下-上)以及彩色编码的各向异性分数(FA)图。为了考虑不同队列间影像采集和参与者特征的差异,研究人员在每个队列中分别制作了感兴趣区域(ROI)模板。为此,随机选取了每个队列中年龄和性别匹配的参与者(UKB中n = 150;ADNI、IMAGEN、ABCD和HCP中n = 100)。在FA图上,研究人员于双侧投射(上部和后部放射冠)和联合(上纵束)纤维处放置了4个直径为4毫米的ROI,这些位置的深髓静脉与脑室体部垂直。三位神经科医生(SYH、YRZ和JD,分别具有4年、7年和8年的经验)在不知情临床数据的情况下独立放置ROI以确保可靠性。这些ROI图被合并为一个4D图,随后被平均化以生成一个单一的3D概率图。我们采用手动可视化来确认配准的准确性和概率图上ROI的位置。研究人员使用不同的概率阈值来选择在脑室水平上最佳覆盖投射和联合纤维的概率图。选定的ROI图被扭曲到MNI152空间,并利用从个体FA扭曲到1 mm FMRIB58_FA模板(空间分辨率,1 × 1 × 1 mm³)过程中得出的估计非线性变形进行二值化,作为模板。沿x轴、y轴、z轴的弥散图以及彩色编码的FA图也被扭曲到MNI空间。研究人员分别记录了投射和联合纤维上四个ROI的x轴(Dx)、y轴(Dy)和z轴(Dz)方向的弥散性,分别记为Dxproj、Dyproj、Dzproj、Dxassoc、Dyassoc、Dzassoc。左右半球的ALPS指数分别计算为
![]()
。平均ALPS指数(mALPS-index)则是双边值的平均值。较高的ALPS指数代表更大的扩散性。在分析之前,研究者在发现队列中排除了患有流行性痴呆、中风、脑积水和颅内肿瘤的个体(流行率:1.5%),以及在ADNI队列中患有流行性痴呆的个体(流行率:10.5%),因为这些疾病可能会影响ALPS指数的估计。并且删除了超出平均值三倍标准差范围的ALPS异常值。在分析之前,ALPS指数通过基于秩的逆正态变换进行了标准化。
5.4 ALPS指数的可重复性
ALPS指数的可重复性在很大程度上受到感兴趣区域(ROI)设置的影响,这些区域应放置在投射纤维区域和联合纤维区域。在ALPS指数的提取过程中,对分割准确性的评估优先考虑了ROI的放置。为了验证使用群体平均ROI模板进行半自动化ALPS指数计算的分割准确性,研究人员评估了从群体平均ROI模板和手动放置的ROI中得出的ALPS指数的可比性。后者经过视觉检查以确保准确性,特别避免在混合皮层下纤维的区域放置ROI。具体而言,研究者从英国生物样本库(UKB)队列中纳入了150名受试者,从IMAGEN队列中纳入了100名受试者,这些受试者最初是随机选择用于生成群体平均ROI模板的。然后,根据扫描时间,将每个队列中的参与者平均分为训练集和测试集。使用上述方法在训练集中生成了平均ROI模板。然后在测试集中分别基于该ROI模板和手动放置的ROI生成了两组ALPS指数。对这两组ALPS指数进行了皮尔逊相关性分析。
5.5 遗传力估计和全基因组关联分析
研究人员使用基于基因组的限制性最大似然(GREML)方法,通过GCTA v1.93.2软件估计了ALPS指数(均值、左侧、右侧)的基于单核苷酸多态性(SNP)的遗传力,同时调整了年龄、年龄的平方、性别、成像中心和前十个遗传主成分(PCs)。利用LDSC v1.0.1软件,基于GWAS汇总数据进行了基于GWAS的遗传力分析。研究人员使用1000基因组计划中欧洲人群的连锁平衡(LD)评分作为参考,并计算了LDSC截距以评估基因组膨胀。
作为主要的GWAS分析,研究人员在英国生物样本库(UKB)中通过PLINK v2.0软件对平均ALPS指数(mALPS-index)进行了GWAS。研究人员应用了线性加性回归模型,并回归了与GCTA分析中相同的协变量(有关基因组风险位点识别、功能注释和FINEMAP的详细信息见补充方法)。鉴于mALPS指数是左右半球ALPS指数的平均值,研究人员分别对左侧和右侧ALPS指数进行了GWAS,作为敏感性分析。为了验证独立显著SNPs的稳健性并排除白质完整性潜在影响,研究人员在英国生物样本库发现队列中将大脑平均各向异性分数(FA)和平均弥散率(MD)作为额外协变量进行了敏感性分析(见补充方法)。主要和敏感性全基因组关联分析的显著性阈值为P < 5.0 × 10^-8。
5.6 验证和泛化
研究人员对主要分析中识别的与ALPS相关的独立显著变异进行了两次验证分析。内部验证在具有白人但非英国血统的参与者中进行(年龄范围 = 45-81岁,分析样本 = 3470)。外部验证在ADNI队列中进行(年龄范围 = 55-75岁,分析样本 = 260)。并且回归了与主要分析中相同的协变量,并使用了相同的分析模型。显著性验证定义为与主要分析一致的效应方向以及经过假发现率(FDR)校正的P值 < 0.05。
为了探究与ALPS相关的独立显著变异是否对年轻个体的ALPS有影响,研究人员在儿童(ABCD,N = 4445)、青少年(IMAGEN,N = 1888)和年轻成年人(HCP,N = 228)中进行了泛化分析。泛化分析使用了与主要分析相同的协变量和关联模型。显著性泛化定义为与主要分析一致的效应方向以及经过假发现率(FDR)校正的P值 < 0.05。
5.7 元回归分析
元回归分析被用来研究年龄依赖性遗传效应对平均ALPS指数(mALPS-index)的影响。研究人员将发现样本按照年龄分为三个三分位数(45-61岁、62-68岁、69-82岁,见表1),并在每个年龄亚组内重新运行与mALPS指数相关的独立显著变异的遗传关联分析。使用R语言包“metafor”,在假设线性和二次关系的情况下,采用逆方差加权固定效应方法,对三个英国生物样本库(UKB)年龄亚组以及ABCD、IMAGEN、HCP和ADNI的遗传关联结果进行元分析。通过假发现率(FDR)校正后P值低于0.05的变异被认为是显著的。
5.8 纵向变化的GWAS
为了评估与mALPS指数横断面变异相关的遗传变异是否也影响其纵向变化,研究人员在两个队列中进行了mALPS相关变异与mALPS指数年变化率之间的遗传关联分析。这两个队列分别是ABCD(基线平均年龄 = 10岁,平均随访时间 = 2.0年,N = 2152)和UKB(基线平均年龄 = 62岁,平均随访时间 = 2.6年,N = 3580,见表1)。研究人员使用R(版本4.2.0)中的Wilcoxon符号秩检验(配对样本Wilcoxon检验)来比较这两个纵向队列中基线和随访时ALPS指数的差异。双侧P < 0.05被认为是具有统计学意义的。纵向变化的GWAS分析模型和协变量与主要分析相同。
5.9 与其他表型的共享遗传变异
研究人员利用GWAS汇总统计数据,通过LDSC v1.0.1软件,使用1000基因组计划中欧洲人群的数据作为参考,估计了ALPS指数与其他相关表型之间的整体遗传相关性。他们进行了全脑范围的LDSC分析,以研究ALPS指数与各种脑表型之间的遗传相关性,这些脑表型包括脑结构(以体积表示)、白质完整性(以各向异性分数(FA)表示)和脑功能表型(以节点振幅和边指标表示)。这些影像表型的选择基于其生物学意义和数据的可用性。并且他们检验了ALPS指数与潜在危险因素(体重指数(BMI)、睡眠、舒张压(DBP)、收缩压(SBP)和脉压(PP)、握力)、认知(反应时间和一般认知功能)、临床诊断(阿尔茨海默病(AD)、注意力缺陷多动障碍(ADHD)、自闭症谱系障碍(ASD)、行为变异型额颞叶痴呆(bvFTD)、中风及其亚型、精神分裂症(SCZ)、双相情感障碍(BP))、脑小血管病(cSVD)的MRI标志物(白质高信号(WMH)、基底神经节、白质和海马中的PVS负荷)以及阿尔茨海默病的脑脊液生物标志物(脑脊液Aβ42、pTau和tau)之间的遗传相关性。经过Bonferroni校正后P值 < 0.05的遗传相关性被认为是显著的。随后,研究人员使用多效性信息条件假发现率(condFDR)分析和双向两样本Mendelian randomization(MR)分析来识别ALPS指数与神经精神和神经系统疾病(AD、ADHD、ASD、bvFTD、中风、SCZ、BP)以及疾病相关生物标志物(WMH、三个PVS测量指标和三个阿尔茨海默病脑脊液生物标志物)之间的遗传重叠和因果关系(详细信息见补充方法)。
精读分享
ALPS指数遗传基础与跨生命周期效应
本研究通过全基因组关联分析(GWAS)揭示脑淋巴系统活动(以DTI-ALPS指数为指标)的遗传机制,首次系统解析其与神经发育及退行性疾病的关联,并探索年龄依赖性遗传效应
优势
a. 基于多队列(UKB、ADNI等)跨年龄段数据(9-82岁,N=40,866),覆盖生命周期;
b. 结合纵向设计与功能注释,揭示ALPS指数动态变化及生物学通路;
c. 发现GMNC、C16orf95等共享遗传位点,链接脑脊液tau病理与脑室体积。
主体内容
研究背景与目标
脑淋巴系统通过清除代谢废物维持脑稳态,其功能障碍与ADHD、自闭症及阿尔茨海默病相关;
探究ALPS指数的遗传架构,解析其与神经表型的多效性及跨年龄遗传异质性
方法与发现
遗传位点:发现17个全基因组显著位点(如12q23.3的NUAK1),定位161个候选基因;
年龄效应:2p23.3位点遗传效应在青年与老年群体中一致,20q13.12位点(SULF2)在衰老中作用增强;
多效性分析:ALPS指数与脑室体积(rg=-0.47)、脑小血管病标志物(WMH)、AD生物标志物(CSF tau)显著遗传相关;
因果推断:孟德尔随机化提示左侧ALPS指数升高可降低ADHD风险(OR=0.647)
挑战与未来展望
ALPS指数与脑淋巴功能直接验证不足,需结合多模态影像与病理研究;
样本未覆盖全生命周期(如胎儿期及中年群体),年龄分层残留混杂;
部分遗传位点未充分复制,需大样本验证及功能实验解析机制
结论
本研究首次绘制ALPS指数的遗传图谱,揭示其与神经退行及精神疾病的共享遗传基础,为脑淋巴靶向干预提供新方向。NUAK1、SULF2等基因的年龄依赖性效应提示生命周期特异性调控机制,未来需结合多组学深化转化研究。
参考文献
Huang SY, Ge YJ, Ren P, Wu BS, Gong W, Du J, Chen SD, Kang JJ, Ma Q, Bokde ALW, Desrivières S, Garavan H, Grigis A, Lemaitre H, Smolka MN, Hohmann S; IMAGEN Consortium; Feng JF, Zhang YR, Cheng W, Yu JT. Genome-wide association study unravels mechanisms of brain glymphatic activity. Nat Commun. 2025 Jan 13;16(1):626.