Rice Science∣武汉大学水稻研究团队发现水稻壁相关激酶OsWAKg16和OsWAKg52同时调控水稻抗病性和产量
近日,农学领域国际期刊Rice Science在线发表了武汉大学杂交水稻全国重点实验室范峰峰博士题为“Identification and Characterization of WAKg Genes Involved in Rice Disease Resistance and Yield”的研究论文。该论文系统分析了水稻壁相关激酶中包含半乳糖醛酸结合结构域的基因家族(WAKg),并评估了其与水稻抗病性和产量的关系。在水稻中共鉴定出108个OsWAKg基因。启动子顺式元件分析显示,这些基因的启动子区域含有丰富的抗性和激素相关元件。进一步分析发现OsWAKg16和OsWAKg52正调控水稻抗病性和产量。此外,OsWAKg52通过多重模式触发免疫(PTI)反应调节水稻抗病性。这些发现表明,WAK基因在水稻抗病性和产量的协调调控中发挥着重要作用,为水稻WAK基因家族研究提供了新的见解,也为作物协同改良提供了潜在的遗传资源。
壁相关激酶(wall-associated kinase, WAKs)是一个在抗病中起着重要作用的基因家族。典型的WAK蛋白包含三个功能域:GUB_WAK_bind(壁相关激酶半乳糖醛酸结合),EGF(表皮生长因子)和PKc_like(蛋白激酶C)蛋白激酶结构域。目前针对WAK基因家族的研究主要集中在EGF相关结构域和蛋白激酶结构域。WAK基因家族的特征是将细胞外基质物理连接到细胞壁上,而GUB_WAK_bind结构域作为直接结合细胞壁的关键功能结构域,却没有得到足够的重视。在这项研究中,作者重点研究了同时包含GUB_WAK_bind结构域和蛋白激酶结构域的WAK蛋白,并将它们归类为壁相关激酶半乳糖醛酸结合(wall-associated kinase galacturonan-binding, WAKg)家族基因。在水稻中共鉴定到108个WAKg基因。随后评估了这些WAKg基因在水稻中的进化关系。结果表明,粳稻与普通野生稻之间有48对共线基因,与籼稻之间有75对共线基因。此外,在普通野生稻、籼稻和粳稻之间鉴定出37对保守共线对(图1-A和-B),这表明WAKg基因在水稻中具有高度保守性。基因重复事件是家族基因形成的必要条件,而串联重复分析显示WAKg家族的扩展不是通过基因复制形成的(图1-C)。
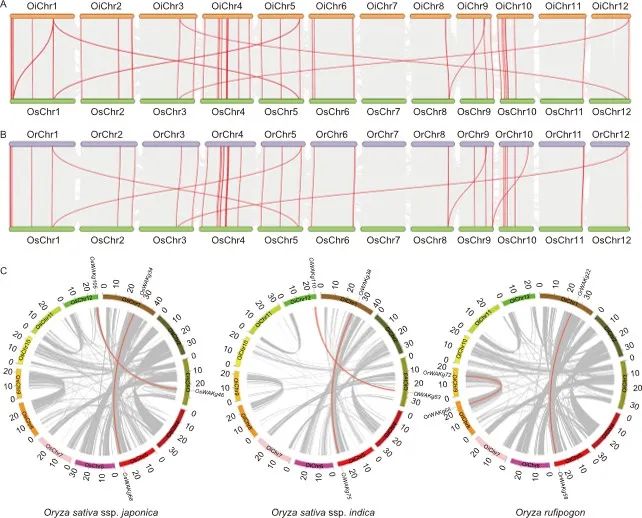
图1. 水稻WAKg家族基因的共线性关系及基因片段重复分析
A和B, 水稻WAKg基因的共线性关系。灰线表示共线块,红线表示WAKg共线性基因对。Oi:籼稻;Os:粳稻;Or:普通野生稻;Chr:染色体。C, 水稻WAKg家族基因片段重复分析。灰色线表示水稻基因组中的所有共线性块,红色线表示重复的WAKg基因对
启动子顺式元件分析显示,OsWAKg基因的启动子区域含有丰富的抗性和激素相关元件。为了进一步研究OsWAKgs与抗性和激素反应之间的关系,作者首先利用RiceXPro网站上的数据集检测了OsWAKgs的时空表达模式,并对其中在叶片和根中表达量最高的一个亚群中的18个基因进行了重点分析。结果显示,这18个基因中,12个基因(OsWAKg10、11、16、23、40、52、53、60、61、70、75和96)受几丁质诱导表达,11个基因(OsWAKg11、13、16、23、40、52、53、60、61、75和96)受稻瘟病菌诱导表达,9个基因(OsWAKg8、10、16、23、40、52、60、70和96)受白叶枯菌诱导表达(图2-A至-C)。这暗示这些基因在免疫应答中起着重要作用,可能有助于水稻抗病反应。随后的激素应答反应显示,6个基因(OsWAKg8、11、13、52、70和96)受吲哚-3-乙酸(IAA)诱导表达,8个基因(OsWAKg8、11、13、16、52、53、70和96)受 BR诱导表达,7个基因(OsWAKg6、8、16、52、53、70和96)受赤霉素(GA3)诱导表达(图2-D至-F)。这些发现表明,这些基因参与激素响应,并可能在水稻生长中发挥作用。在所有分析的基因中,8个基因(OsWAKg8、11、13、16、52、53、70和96)对病原体和激素均有应答(图2),暗示这些基因可能在水稻抗病和生长调节中具有双重作用。
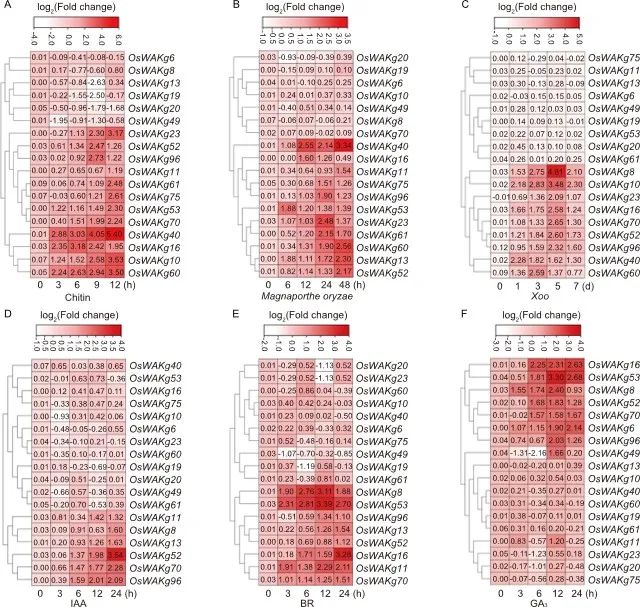
图2. 几丁质(A)、稻瘟病菌(B)、白叶枯病菌(C)、吲哚-3-乙酸(D)、油菜素内酯(E)和赤霉素(F)诱导表达分析
对叶和根中表达最高的18个OsWAKgs进行分析。10日龄的幼苗进行几丁质诱导,12日龄的幼苗进行激素(IAA、BR、GA3)诱导,30日龄的叶片进行稻瘟病菌(菌株GY1173)和白叶枯病菌(菌株PXO99)诱导
为了验证上述可能在水稻抗病和生长调节中具有双重作用基因的功能,对其进行了敲除转基因验证。OsWAKg16敲除系(oswakg16-1和oswakg16-2)表现出缺失突变,导致蛋白质翻译提前终止(图3-A)。抗性分析显示,oswakg16与野生型相比,病斑长度和真菌生长速度菌显著增加(图3-B至-E)。这说明oswakg16的稻瘟病抗性显著下降。在农艺性状方面,oswakg16的粒长变化不显著,但粒宽明显减小,千粒重显著降低(图3-F至-J);此外,oswakg16的株高和每穗粒数显著降低,而有效穗数和结实率保持不变(图3-K至-M);这些变化最终导致其单株籽粒产量下降24.2%左右(图3-N)。上述结果表明,OsWAKg16对水稻稻瘟病抗性和产量均有正向调控作用。
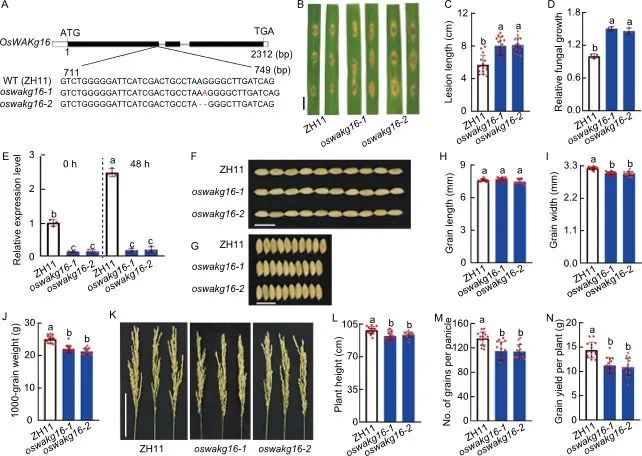
图3. OsWAKg16的功能分析
A, OsWAKg16敲除系(oswakg16-1和oswakg16-2)中的突变位点示意图,以中花11(ZH11)作为野生型(WT);B, 稻瘟病抗性分析,用稻瘟病菌分离物GY1173对30日龄叶片进行接种,接种后6天拍摄叶片;C, 在接种6 天后测量相对病变面积;D, 在接种6 天后测量真菌的相对生长;E, 稻瘟病菌接种前后的OsWAKg52表达量分析;F-J, 野生型和oswakg16的粒长、粒宽和千粒重表型;K-N, 野生型和oswakg16的穗型、株高、穗粒数和单株产量表型
另一个基因OsWAKg52的敲除系(oswakg52-1和oswakg52-2)表现出插入或缺失突变,均导致提前终止(图4-A)。抗性分析显示,稻瘟病菌接种后,oswakg52表现出明显增加的病斑形成和真菌生长速度(图4-B至-E);同样的,白叶枯病菌接种后,oswakg52显示病斑形成和细菌生长显著增加(图4-F至-I)。与此同时,水稻叶片中OsWAKg52的表达在稻瘟病菌、白叶枯病菌或几丁质处理后显著升高(图4-E和-I)。这些结果表明OsWAKg52是水稻抗病的正向调节因子。在农艺性状方面,oswakg52的粒长明显增加,粒宽明显减小,长宽比明显增加,但千粒重明显降低(图4-J至-O)。在其他农艺性状不变的情况下,千粒重的减少直接导致了产量的大幅下降(图4-P)。这些结果与抗病试验相结合,表明OsWAKg52正调控水稻抗病性和产量。
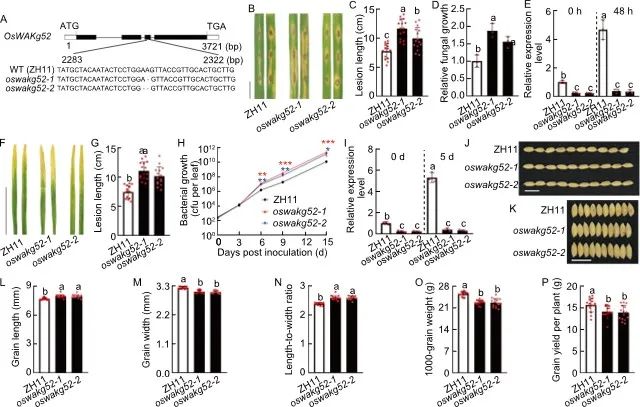
图4. OsWAKg52的功能分析
A, OsWAKg52敲除系(oswakg52-1和oswakg52-2)中的突变位点示意图。以中花11(ZH11)作为野生型(WT);B, 稻瘟病抗性分析,用稻瘟病2分离物GY1173对30日龄叶片进行接种,接种后6 d拍摄叶片;C, 在接种6 天后测量相对病变面积;D, 在接种6 天后测量真菌的相对生长;E,稻瘟病菌接种前后的OsWAKg52表达量分析;F, 白叶枯抗性分析,用白叶枯病菌PXO99对30日龄叶片进行接种,接种后14天拍摄叶片;G, 在接种14天后测量病变长度;H, 在接种14天后测量细菌的相对生长;I, 白叶枯病菌接种前后的OsWAKg52表达量分析;J-P, 野生型和oswakg16的粒长、粒宽、长宽比、千粒重和单株产量表型
为了进一步探究OsWAKg52介导的抗病分子机制,作者分析了其生物学特性。qRT-PCR分析显示,OsWAKg52在所有组织中均有表达,但在叶片中表达量最高。OsWAKg52-GFP融合蛋白定位于细胞核、细胞质和细胞膜,表明OsWAKg52是一个泛定位蛋白(图5-A)。随后,作者选择了抗性较弱的oswakg52-1系测量几丁质处理下活性氧(ROS)爆发和防御相关基因表达。与野生型相比,oswakg52-1在几丁质处理下ROS爆发明显减少(图5-B)。在几丁质处理下(图5-C至-F)和稻瘟病菌感染期间(图5-G至-J),oswakg52-1中防御相关基因(pid2、PR3、PR5-1和PR10)的表达均显著下调。稻瘟病菌侵染后,oswakg52-1的过氧化氢和超氧阴离子水平也显著降低(图5-K和-L)。与此同时,DAB(二氨基联苯胺)和NBT(硝基蓝四氮唑)染色证实oswakg52-1中ROS积累减少(图5-M)。综述所述,OsWAKg52通过调控多重PTI反应来调控水稻对稻瘟病和白叶枯病的抗性。
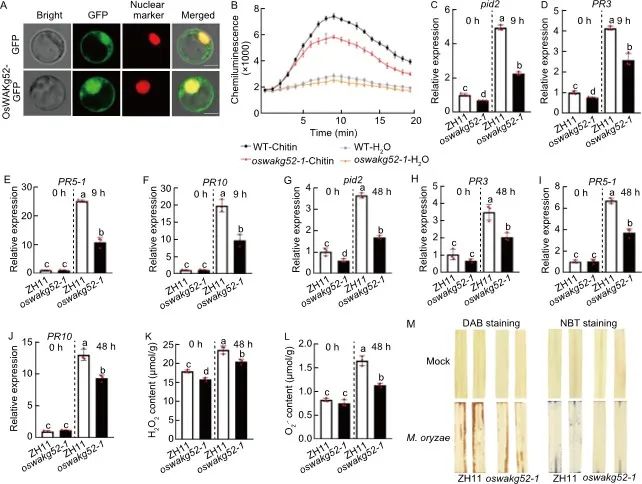
图5. OsWAKg52通过正向调节PTI增强对水稻稻瘟病和白叶枯病的抗性
A,OsWAKg52-GFP融合蛋白在水稻原生质体中的亚细胞定位;B,几丁质诱导后活性氧(ROS)积累检测;C-F,几丁质诱导前后野生型和oswakg52-1中Pid2 (C)、PR3 (D)、PR5-1 (E)、PR10 (F)的相对表达量,使用10天幼苗的叶片进行诱导处理;G-J,稻瘟病菌诱导前后Pid2 (G)、PR3 (H)、PR5-1 (I)、PR10 (J)在野生型和oswakg52-1中的相对表达量,使用30天幼苗的叶片进行诱导处理;K和L,分别测定稻瘟病菌侵染48小时后野生型和oswakg52-1叶片中过氧化氢 (K)和活性氧含量(L);M,稻瘟病菌侵染48小时后野生型和oswakg52-1叶片的DAB(二氨基联苯胺)和NBT(硝基蓝四氮唑)染色
本研究明确了WAKg与水稻抗病和生长发育密切相关,并鉴定出OsWAKg16和OsWAKg52是水稻抗病和产量的正调控因子。这些发现为水稻WAK基因家族提供了新的认识,并为协同提高水稻抗病性和粮食产量提供了潜在的遗传资源。
武汉大学杂交水稻全国重点实验室的博士生Ayaz Ahmad和武汉生物工程学院的青年教师程明星为共同第一作者,武汉大学杂交水稻全国重点实验室的李绍清教授和范峰峰博士为共同通讯作者。该研究得到了国家自然科学基金面上项目(32470391)及青年项目(32401801),崖州湾实验室与中国种子有限公司联合揭榜挂帅项目(B23YQ1515),湖北省国际合作项目(2023EHA045)和武汉生物工程学院高层次人才科研启动基金项目(2024KQ07)等项目支持。
论文链接:
http://www.ricesci.org/EN/abstract/abstract10305.shtml